

Effects of nutrition on feathering

Diet nutrient profile, feed intake/restriction, anti-nutrients and various feed additives can all influence feather development from early age onwards. Because of the high protein and amino acid content of feathers, these nutrients deserve the most attention.
Early studies using a simple feather litter score system, showed that crude protein content of the diet had a direct effect on feather regeneration. More feathers were found on the litter when a high protein diet was fed, being indicative of greater feather growth and development and subsequent juvenile moult. On the other hand, absence of feathers on the litter is sometimes cause for concern. Deficiency of crude protein is known to have the opposite effect, where poorly feathered chicks will invariably be seen if birds younger than 10-15 d of age are given diets with much less than 16% crude protein, regardless of diet amino acid profile. This situation suggests that either there is a requirement for protein per se for feathers, or that we have not correctly established the optimum balance of amino acids needed for feather growth independent of other traits such as growth and feed efficiency. However, its not just balance of amino acids per se that impact feathering, rather its the intake of this balanced ratio. The impact of unintentional feed restriction is undoubtedly a factor at high stocking densities in the later part of the broiler growing cycle.
Amino acids
Any discussion on the involvement of nutrition on feathering invariably focuses on the cystine and associated methionine level of the diet, and the first reaction is to add methionine to the diet. The major amino acids involved in the synthesis of feather keratin are the sulphur containing amino acids, cystine and methionine. The relative proportion of the sulphur containing amino acids is much greater in the integument than in muscle tissue and so marginal dietary deficiencies of these amino acids will often be initially manifested as abnormal feathering. Cystine is the most common amino acid in feathers, representing some 9% of total amino acids, although valine, arginine and leucine are also in high concentration. It is an interesting exercise to calculate the proportion of amino acids consumed by a broiler each week that end up in feathers (Table 1). If these calculations are correct, then it indicates that valine and arginine intake are also critical for feather development.
Abnormalities
Another issue often raised when feather abnormalities occur is the source of methionine used, as either DL Methionine or its hydroxy analogue. A review of the literature suggests no difference in these sources in terms of general metabolism and especially for feathering. By using radio-labelled sources, it is obvious that HMB does end up in the feather follicles.
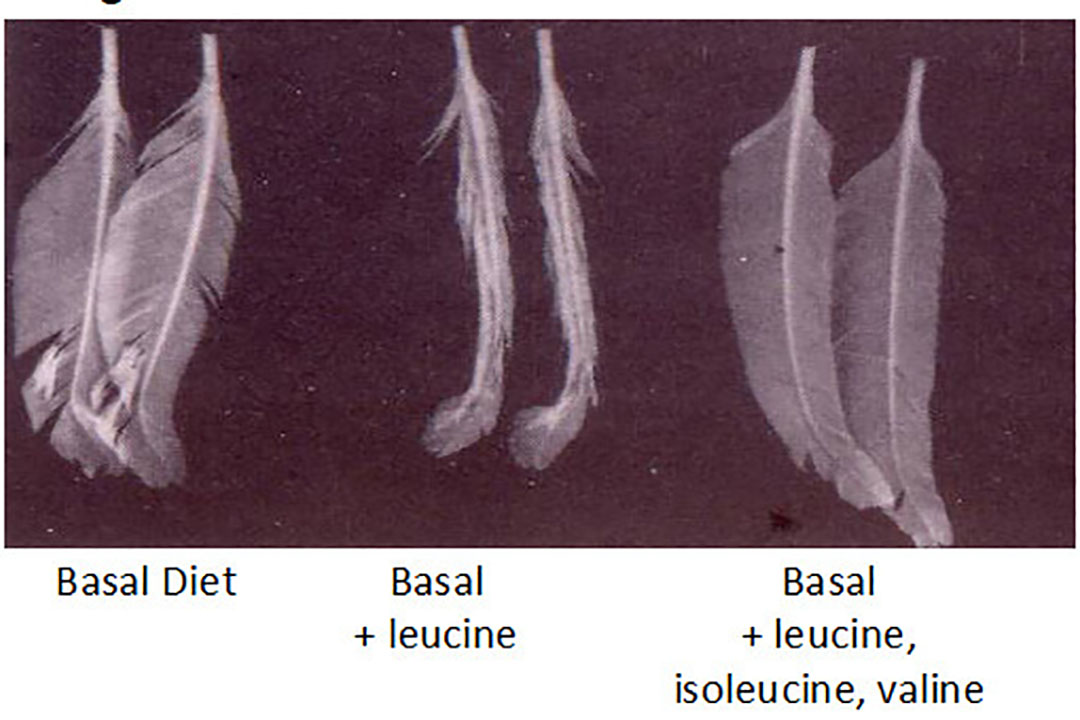
Although abnormal feathering can be used as an aid in diagnosing amino acid and other nutritional deficiencies, results are often non-specific, such that antagonisms, deficiencies and/or imbalances of amino acids (and various other nutrients) often result in similar gross abnormalities of the plumage. For example, imbalance of leucine and valine, or even glycine, phenylalanine and tyrosine both cause similar types of feather abnormalities in young chicks. In most situations, the appearance of affected chicks on such deficient or imbalanced diets is best described as ‘rough feathering’ and upon closer examination the body feathers are often sticking out from the body while primary wing feathers are invariably unsheathed closer to the body. Classical work by Mario Penz some years ago, showed the effect of an imbalance of the branch- chained amino acids of growth of broiler wing feathers (Illustration 1). Adding an excess of leucine caused abnormal feathering, which was corrected by adding a corresponding excess of leucine and isoleucine.
Feather pigmentation is also influenced by nutrition. Lysine has been identified as the major amino acid involved in pigment formation although reduced pigmentation is also seen in chicks fed diets deficient in phenylalanine and tyrosine, which are precursors of melanin. Lysine deficiency may also cause depigmentation by inhibiting activity of the enzyme tyrosinase so inhibiting the formation of melanin.
Vitamins and minerals
Deficiency of most vitamins and minerals cause abnormal feathering, although effects are rarely different enough to allow diagnosis. Deficiency of riboflavin causes perhaps the most characteristic signs of deficiency in embryos and chicks. Affected embryos exhibit clubbed down, which, as its name implies, involves a swollen tip for each down feather (Illustration 2).

In the last few years, there has been increased incidence of clubbed down in chicks, visible at the hatchery. This is sometimes called defective-down syndrome. The current condition is not responsive to additional riboflavin in the breeder feed. A recent study suggested the cause as an induced zinc deficiency in breeders caused by feeding too high levels of calcium and iron. On the other hand, an astrovirus may be implicated in breeders. There is also anecdotal evidence that clubbed down occurs more frequently in floor vs nest eggs. Many years ago, researchers in Australia suggested that clubbed down associated with so called “ginger hairs” on the neck region of newly hatched chicks, was caused by a metabolic deficiency of molybdenum.

Trace minerals
Inadequate levels of trace minerals will also impede feather development. Deficiencies of zinc, tin, vanadium, chromium and nickel have all been reported to affect feathering. Zinc deficiency has probably been most studied where a deficiency causes frayed feathers especially in the rapidly growing primary wing feathers. Characteristic blisters appear on the shaft of the feathers about 3 cm from the follicle and the feather often breaks at this point. Zinc deficiency also causes “spoon-shaped” feathers, where there are no vanes developing close to the follicle. Spoon-shaped feathers have also been described for birds infected with reticulo-endothelial virus as well as during transient starvation.
Another mineral of current concern is boron toxicity, which is sometimes used as a litter treatment to control darkling beetle. It is not clear if the boric acid causes problems due to contact with the skin, or if birds are affected due to eating the contaminated litter. Iodine is also a critical nutrient for feathering because of the intimate association with thyroid metabolism. In the past, we have likely underestimated the birds need for Iodine re feathering, since for most classes of bird, dietary requirements were invariably published in breeder manuals at 1ppm. Today we realise that 2-3ppm is probably more relevant for optimum feather growth. Corn and soybean meal both contain around 0.2ppm iodine. Wheat is much lower at 0.1ppm while sorghum and millet are exceptional low at 0.03ppm.
Mycotoxins
Dietary mycotoxins are also known to cause feather abnormalities. Birds fed high levels of T-2 toxin develop only a sparse covering of feathers, which tend to protrude from the bird at odd angles. One possibility is that T-2 toxin alters the metabolism of certain nutrients and so effects are indirect. With T-2 toxicosis most of the body feathers are influenced, unlike the localised situation seen with vitamin or amino acid imbalance. However, feather problems occur with around 5-15ppm T-2 toxin in the diet which needs abnormally high levels in corn and that severity is dose related. The feather shafts tend to have an accentuated curve away from the body and in some cases there is a distinct dark bar across the feather about one-quarter the length down from the tip. Ochratoxin has been reported to cause delayed feathering in broilers. It is likely that presence of multiple mycotoxins, at levels that are singularly not important, can have adverse effects on feathering even when all nutritional requirements and levels are met.





